����ʬ�ͻï�
���A�S�٬��ï��A�O���㦳�ʤ��\��J�ս��C[1]�b�����ʤƤ�����t���A���������l�Q�٬������A����q�L�����ʤ���Ƭ��t�@�ؤ��l�C�X�G�Ҧ����ӭM���ʶi�{���ݭn�����ѻP�A�H�����IJv�C�P��L�D�ͪ��ʤƾ��ۦ��A���q�L���C�ƾǤ��������Ư��]��Ea�ΣGG�ު��ܡ^�ӥ[�֤����t�v�A�j�h�ƪ����i�H�N��ʤƪ��������t�v�����W�ʸU���F�P�ˡA���@���ʤƾ��A�����b�����L�{�����Q���ӡA�]���v�T�������ƾǥ����C�P��L�D�ͪ��ʤƾ����P���O�A���㦳���ת��M�@�ʡA�u�ʤƯS�w�������β��ͯS�w���c���C�ثe�w�����i�H�Q���ʤƪ���������4000�ءC[2]
���M���O�J�ս�A���ëD�㦳�ͪ��ʤƥ\����l���O�J�ս�A���@�dzQ�٬��֖���RNA���l�P�˨㦳�ʤƥ\��C[3]���~�A�q�L�H�u�X���ҿ��H�u���]�㦳�P���������ʤƬ��ʡC[4]���H�{�������w�q���㦳�ʤ��\��ͪ��j���l�A�Y�ͪ��ʤƾ��A�h�өw�q�����]�t�㦳�ʤƥ\��J�ս�M�֖��C[5]
�����ʤƬ��ʥi�H����L���l�v�T�G����O�i�H���C�����ʪ����l�F���ƾ��h�O�i�H�W�[�����ʪ����l�C���\�h�Ī��M�r���N�O��������C���������٥i�H�Q�ū��B�ƾ����ҡ]�ppH�ȡ^�B����@�ץH�ιq�Ϫi�]�p�L�i[6]�^���\�h�]���Ҽv�T�C
���b�u�~�M�H�̪���`�ͬ��������Τ]�D�`�s�x�C�Ҧp�A�ļt�ίS�w���X�����ӦX���ܥͯ��F�[���~�篻�q�L���ѳJ�ս�M�ת������U���h�窫�W���ú{�M�o�{�C
�o�{�ά�s�v
�����o�{�ӷ���H�̹��o�����z���v���F�ѡC���b18�@�����M19�@����A�H�̴N�{�Ѩ쭹���b�G���Q�����A[7]�δӪ��������G�i�H�N������Ƭ��}�A��������������z�h�ä��F�ѡC[8]
��F19�@�������A�k���Ǯa�����P�ڴ��w�����}��Ƭ��s�����o�ùL�{�i��F��s�A�{���b�å��ӭM���s�b�@�ج��O����A�R�W���u�ï��v�]ferment�^�C�L���X�o�ìO�o�ج��O����ʤƪ����G�A�û{�����O����u�s�b��ͩR�餤�A�ӭM�}���N�|���h�o�ç@�ΡC[9]
1878�~�A�w���Ͳz�Ǯa�·G�P�}���������X�F���]enzyme�^�o�@�����C�H��A���Q�Ω�M���G�J�Ֆ����@���D���骫��A���ï��]ferment�^�h�Q�Ω���Ѭ���ӭM���ͪ��ʤƬ��ʡC
�o�ع�����~�{�ѫܧֱo��ȥ��C1897�~�A�w���Ǯa�R�w�ءP��Ư��}�l�藍�t�ӭM���å������G�i��o�ì�s�A�q�L�b�f�L�x���j���Ұ����@�t�C����̲��ҩ��o�ùL�{�ä��ݭn���㪺���ӭM�s�b�C[10]�L�N�䤤����o���o�ç@�Ϊ����R�W���o�Ö��]zymase�^�C[11]�o�@�^�m���}�F�q�V�{�N�����P�{�N�ͪ��ƾ����j���A�䥻�H�]�]�u�o�{�L�ӭM�o�äά������ͤƬ�s�v����o�F1907�~���ը����ƾǼ��C�b������A���M�ï���ӷ����X�G���@�A�è̾ڤ�ƯǪ��R�W��k�A�����o�{�̭̮ھڨ�Ҷʤƪ������N���̩R�W�C�q�`�����^��W�٬O�b�ʤƨ���Ϊ̤����������W�r�̫�[�W-ase�����A�ӹ�������R�W�]�ĥ�������k�A�Y�b�W�r�̫�[�W�u���v�C�Ҧp�A�ſ}���]lactase�^�O����Ť��ſ}�]lactose�^�����FDNA�E�X���]DNA polymerase�^����ʤ�DNA�E�X�����C
�H�̦b�{�Ѩ얡�O�@�����̿����ӭM�������A�U�@�B�u�@�N�Oų�w��ͤƲզ������C�\�h������s�̫��X�A�@�dzJ�ս�P�����ʤƬ��ʬ����F���]�A�ը������o�D���L�w�P�����I���S�b����������Ǯa�{�������O�J�ս�A�L���G�٨��dzJ�ս�u�O�����l����a�̡A�J�ս襻���ä��㦳�ʤƬ��ʡC1926�~�A�����ͪ��ƾǮa��i���P�ĩi�������F�@�ӨM�w�ʪ�����C�L�����q�M���o�������������A���ҩ��F���������J�ս�����C���A�ĩi�Ǧb1931�~�b�L��ƲB������s���A���ҹ�F�����J�ս�C�����P�N�ؼw�P�ի�ù���M�żw���P���p�}��P���Z�Q�q�L��G�J�Ֆ��B�سJ�Ֆ��M�ؾ��ųJ�Ֆ������Ʃ��J�Ֆ�����s�A�̲T�{�J�ս�i�H�O���C�H�ᳰ��o�{����d�E�ؖ����ҩ������ƾǥ���O�J�ս�C�H�W�T���Ǯa�]����o1946�~�ը����ƾǼ��C[12]
�ѩ�J�ս�i�H�����A�q�LX�g�u������N�i�H����T�����c�i���s�C�Ĥ@����o���c�ѪR�������l�O���ߖ��A�@�ئb���\�B��G�M�J�M���t�q�״I�����A��\��O�����ӵ��~�ߡC���ߖ����c���j�áP��Q���]David Phillips�^�һ�ɪ���s�ոѪR�A�é�1965�~�o���C[13]�o�@���G���o���Чӵ����c�ͪ�����s���}�l�A���ѪR�ת����T�����c�ϱo��b���l�����W���u�@����F�Ѧ����i��C
1980�~�N�A�������P�����]Thomas Cech�^�M�����P�����S�����O�q�|������rRNA�e�骺�[�u��s�M�ӵߪ��ֿ}�ֻĖ�P�ƦX������s�����o�{RNA�����㦳�ۧڶʤƧ@�ΡA�ô��X�F�֖��������C�o�O�Ĥ@���o�{�J�ս�H�~���㦳�ʤƬ��ʪ��ͪ����l�C 1989�~�A��G�H�]�]����o�ը����ƾǼ��C[14]
[�s��] �ͪ��ǥ\��
�b�ͪ��餺�A���o���۫D�`�s�x���\��C�H������M�ӭM���ʪ��ձ��������}���A�S�O�O�E���M�C�Ė����ѻP�C[15]���]�ಣ�B�ʡA�q�L�ʤ��ٲy�J���WATP�����Ѳ����٦צ��Y�A�åB����@���ӭM���[���@�����ѻP�B�e�M������C[16]�@�Ǧ���ӭM���W��ATP���@�����l���ѻP�D�ʹB���C�@�ǥͪ��餤����_�S���\��]�������ѻP�A�Ҧp���������i�H���ä����o���C[17]�f�r���]�t�����A�ΰѻP�I�V�ӭM�]�pHIV��X���M�f������^�A�ΰѻP�f�r���ɱq�J�D�ӭM������]�p�y�P�f�r�����g�i�Ė��^�C
�����@�ӫD�`���n���\��O�ѻP�b�ʪ����ƨt�����u�@�C�H�������M�J�Ֆ����N�����@�ǖ��i�H�N�i�J���ƹD���j���l�]�����M�J�ս��^���Ѭ��p���l�A�H�K��z�D�l���C��������Q�z�D�����l���A�Ӗ��i�H�N�������Ѭ����}�Χ�i�@�B���Ѭ�����}���z�D�i�H�l�����p���l�C���P�������Ѥ��P�����������C�b���ϯ�ʪ������ƨt�Τ��s�b�@�ǥi�H�����ֺ��������ӵߡA�ֺ������i�H���ѴӪ��ӭM�������ֺ����A�q�Ӵ��ѥi�Q�l�����i�ơC
�b�N�³~�|���A �h�Ӗ��H�S�w�����ǵo���\��G�e�@�Ӗ��������O��@�Ӗ�������F�C�Ӗ��ʤƤ�����A�����Q�ǻ���t�@�Ӗ��C���DZ��p�U�A���P�����i�H����a�ʤƦP�@�Ӥ� ���A�q�Ӥ��\�i��������ձ��G��p�@�Ӗ��i�H�H���C�����ʫ���a�ʤƸӤ����A�ӥt�@�Ӗ��b�Q���ɫ�i�H���������ʶi��ʤơC�����s�b�T�w�F��ӥN�«� ���T���~�|�i��F�Ӥ@���S�������s�b�A�N�¬J������һݨB�J�i��A�]�L�k�H�������t�ק����X���H�����ӭM���ݭn�C��ڤW�p�G�S�����A�N�³~�|�A�p�}�ø��A�L�k�W�߶i��C�Ҧp�A����}�i�H�����PATP�����ϱo��@�өΦh�Ӻҭ�l�Q�C�Ĥ��F�b�S�������ʤƮɡA�o�Ӥ����i��o�D�`�w�C�H�P�i�H�����F�Ӥ@���[�J���ҿ}�E���A�b6��W���ҭ�l���C�ĤƤ�����o���j�[�t�A���M��L�ҭ�l���C�ĤƤ����]�b�w�C�i��A���b�@�q�ɶ����˴��i�H�o�{�A���j�h�Ʋ���������}-6-�C���C��O�C�ӲӭM�N�i�H�q�L�o�ˤ@�M�\��ʖ��ӧ����N�³~�|����Ӥ��������C
[�s��] ���c�P�ʤƾ��z
�@���J�ս�A���P�ؖ��������j�p�t�O�D�`�j�A�q62���i����ݰ�4-���Q�ڨ���ܲ��c���]4-oxalocrotonate tautomerase�^[18]��W�L2500�Ӵݰʪ��תջĦX����[19]�C�����T�����c�M�w�F���̪��ʤƬ��ʩM���z�C[20]�j�h�ƪ������n�̪��ʤƨ���j�o�h�A�åB�����l���u���@�p�����]3-4�Ӵݰ�^�������O�ʤƤ����C[21]�o�ǰѻP�ʤƴݰ�[�W�ѻP���X���誺�ݰ�@�P�Φ��F�o�ͶʤƤ������ϰ�A�o�@�ϰ�N�Q�٬��u���ʤ��ߡv�Ρu���ʦ��I�v�C���\�h���t��������X��ʤƤ����ҥ��ݪ����]�l�����X�ϰ�C���~�A�٦��@�ǖ�������X�ʤƤ��������������������Ϊ̨���F�o�ص��X����W�[�έ��C�����A�O�@�����X�ո`��q�C
�P��L�D���J�լۦ��A������P�|�Φ��h�ؤT�����c�����C���@�������O�Ѧh���Ȱ��Ҳզ����ƦX�����C���F�ݼ����������H�~�A�j�h�Ɩ��b���ű��p�U�|�o�ͥh�P�|�A��T�����c�M�����ʳQ�}�a�F��P�����A�o�إh�P�|���i�f�ʤ]���Ҥ��P�C
[�s��] �M�@��

�T�ؖ��ʤƾ���Ҧ��ϡGA. �u��-�_�͡v�Ҧ��FB. ���ɫ��X�Ҧ��F C. �s�鲾�ʼҦ��C
�q�`���p�U�A������Ҷʤƪ����������M��������㦳���ת��M�@�ʡC�������ʦ��I�M����A���̪��Ϊ��B�����q���B�����������|�v�T�M�@�ʡC�����ʤƥi�H�㦳�ܰ�������M�@���B�ϰ��ܩ��M�ƾǿ�ܩ��]chemoselectivity�^�C[22] ����ӻ��A���u��㦳�S�w�Ŷ����c���Y�ةάY������_�@�ΡC�Ҧp�A���}���u����\-����}�����_���ӹ��]-����}�����L�v�T�C���~�A���㦳�������M���c�����ѧO��O�A�u���@����M���@�ΡA�ӹ�t�@��M�餣�_�@�ΡC�Ҧp�A�سJ�Ֆ��u����ѥ�L-�i����Φ��������A�Ӥ���@�Ω�D-�i����Φ�������F�å��������u���D-�c���}(�pD-����})�o�áA�ӹ�L-�c���L�ġC
���P���������M�@�ʮt�O�ܤj�C�@�ǖ�����ѻP�ݭn�������ǽT�ת���]���ƻs�M���F���A�o�ǖ����㦳�u�չ�v����C�HDNA�E�X�����ҡA������������ʤƤ����A�M��A�˴������O�_���T�C[23]�o�ˤ@�رa���չ諸�X������A�ϱo�㦳���O�u�ת����Űʪ��E�X���������X���X�v�C��@�ʸU�����@�A�Y�����@�ʸU�Ӥ����A�X�{�������~����������@�ӡC[24]�bRNA�E�X��[25]�B���QtRNA�X����[26] �M�ֿ}��[27]���]�o�{�F�������չ����C�ӹ��t�@�ǰѻP�X�����ͥN�²����]secondary metabolite�^�����A���̯���P�۹���s�����P����@�ΡC���H�{���o�اC�M�@�ʥi����s���ͪ��X���~�|���i���Q�����n�C[28]
���F���������M�@�ʡA��s�̴��X�F�h�إi����P���誺���X�Ҧ��]���ؼҦ����j�h�Ƭ�s�̩ҶɦV�^�G
[�s��] �u��-�_�͡v�Ҧ��]�uLock and key�v�^
�ӼҦ��������ҡP�J�̺��P�O�����_1894�~���X�A��z�O���M���賣���@�w���~�ΡA�Y�B�߭Y��̤������~�ί����T���ɮɡA�ʤƤ����~�i�H�o�͡C[29]�o�@�Ҧ��q�`�Q�ζH�a�٬��u��-�_�͡v�Ҧ��C���M�o�@�Ҧ�������������M�@�ʡA���o�L�k�����������í�w�������L��A�C
[�s��] ���ɫ��X�Ҧ��]Induced fit�^

���ҿ}�E���b���X ����}���l�e��ҵo�ͪ����c�ܤơC���䬰���X����}���l�e�A�b�Y�ҫ������ʦ��I�F�k�䬰���X��C
�ӼҦ����������P�줰���]Daniel Koshland�^�q�L�ק�u��-�_�͡v�Ҧ��A�_1958�~���X�C��z�O�A�J�M���@���J�ս�A�䵲�c�O�㦳�@�w�X�ʪ��A�]�����ʦ��I�b���X���誺�L�{���A�q�L�P������l�������ۤ��@�ΡA�i�H���_�o�ͷL�p�����ܡC[30]�b�o�@�Ҧ����A���褣�O²��a���X���ʪ����ʦ��I�W�A���ʦ��I�W���i��Ĵݰ������i�H�\�ʨ쥿�T����m�A�ϱo������i��ʤƤ����C�b���X�L�{���A���ʦ��I���_�a�o���ܤơA������觹�����X�A���ɬ��ʦ��I���Ϊ��M�a�q���p�~�|�̲T�w�U�ӡC[31]�b�@�DZ��p�U�A����b�i�J���ʤ��߮ɤ]�O�|�o�ͷL�p���ܪ��A�p�}�������ʤƤ����C[32]
[�s��] �s�鲾�ʼҦ��]Population shift�^
�o�@�Ҧ��O��~�Ӵ��X���@�طs�����P���誺���X�Ҧ��A[33]�� �ϸ����b�@�ǖ����ҵo�{�����赲�X�e��A�����c�H�����j�ܤơA�ӳo�O�λ��ɫ��X�Ҧ��L�k�������C�����]�O�A���b���G���P�ɦs�b���P�c�H�A�@�غc�H �]�c�HA�^���A�X���赲�X���c�H�A�ӥt�@�ء]�c�HB�^�h���A�X�A�o��غc�H�����O���۰ʺA���šC�b�S������s�b�����p�U�A�c�HB���D�ɦa��F���[�J���� ��A�H�ۨ��褣�_�P�c�HA���X�A���G���c�HA�t�q�U���A��غc�H���������ųQ���}�A�ɭP�c�HB���_�a��Ƭ��c�HA�C
���ʤƾ��z�h�ئh�ˡA���~�P�k���O�̲׳�������C�������GG�ޡG[34]
- �гyí�w�L��A���L�����C�Ҧp�A�q�L�P�������L��A���l���˩M�O�]�P������l�ۤ�^�A������í�w�ʡF�Χᦱ������l�A�H�ϱo������ͦV�_��Ƭ��L��A�C
- ���Ѥ��P�������~�|�C�Ҧp�A�Ȯɩʦa�E������A�Φ���-����ƦX���������A�C
- �N���������P������l���X��@�_�A�éT�w����ܤ���������T�o�ͪ���m�A�q�ӭ��C�������u���e�v�C�p�G�u�Ҽ{�������U�ܡ]�GH�ޡ^�A�h���@�η|�Q�����C���쪺�O�A�o�@�@�ΦP�ɤ]�|���C������A��í�w�ʡA[35]�]�����ʤƪ��^�m���p�C[36]
[�s��] �L��A��í�w
���P�@�����b�����ʤƩM�����ʤƪ����p�A�i�H�F�і��O�p��í�w�L��A���C�̦��Ī�í�w�覡�O�q���ۤ��@�ΡA���i�H���L��A���l�W���q�����ѩT�w���ۤϹq���A[37]�ӳo�O�b�����G�D�ʤƤ�����t�����s�b���C
[�s��] �ʺA�@��
�̪@�Ǭ�s���ܤF���������ʺA�@�λP��ʤƾ�������pô�C[38][39]���������ʺA�@�Υi�H�y�z���䤺���զ�����]�p���p�@���i��ġB�@���i��ġF�j���p�@�q���ϰ�B�@���\�����ά۾F���]���F�Ϊ̥i�H�O������c���^���B�ʡA�o�عB�ʥi�H�o�ͦb�q�����]10-15���^��������P�ɶ��ثסC�q�L�o�ذʺA�@�ΡA��Ӗ����l���c�����i��Ĵݰ�N���i�H��ʤƧ@�άI�[�v�T�C[40][41][42][43]�J�ս�ʺA�@�Φb�\�h�������_������@�ΡA�ӬO�p���ֳt�B���٬O�j���۹���C���B�ʰ_�@�Χ�h�O�̿�Ҷʤƪ����������C���ʺA�@�Ϊ��o�Ƿs�o�{�A���F������@���B�]�p�H�u���M�}�o�s�ij������n�N�q�C
���������X���O�A�o�خɶ��̿�ʺA�i�{���j�i�����U�������ʤƤ������t�v�A�]���o�عB�ʬO�H���o�ͪ��A�åB�t�v�`�ƨ��M���F�����A���X�v�]P�^�]P = exp {�GG��/RT}�^�C[44]�ӥB�A���C�GG�ݭn�۹���p���B�ʡ]�P�b���G�������������B�ʬۤ�^�H�F��������P�����������L��A�C�]���A�o�عB�ʩΪ̻��ʺA�@�ι��ʤƤ�������^�m�٤��M���C
[�s��] ����ո`
-
�b���X�����l�����p�U�A���얡������ܦۨ����c�A�q�ӹF��ո`�����ʪ������C�o�ؽո`�@�Υi�H�O�������A�Y�����l���X�첧�얡�W�F�]�i�H�O�������A�Y�����l�q�L���X�䥦����P���얡�ۤ��@�Ϊ��J�ըӵo���ո`�@�ΡC
�@�뱡�p�U�A���b�`�šB�`���M���ʤ����G����U�i�H���`�o���ʤƬ��ʡC�b���ݱ���U�A�]�A���šB�L���ιL�CpH���A���|���h�ʤƬ��ʡA�o�Q�٬����������C���]���@�ǖ��h���n�b�D�`����U�o���ʤƥ\��A�p�ݼ����������b���ű���U�ϦӨ㦳�������ʡA�ݻĵ��������S���n�CpH����C
[�s��] ���]�l�P����
-
[�s��] ���]�l
�ëD�Ҧ������ۨ��N�i�H�ʤƤ����A���@�ǖ��ݭn���X�@�ǫD�J�դp���l��~�i�H�o���δ����ʤƬ��ʡC[45]�o�Ǥp���l�Q�٬����]�l�A���̬J�i�H�O�L�����l�����l�]�p�������l�B�K���L�^�A�]�i�H�O�����ƦX���]�p�����B������^�C�������]�l�q�`�O�����A�i�H�P����������D�`�c�T�a���X�C�o�بc�T���X�����]�l�P�����]�pNADH�^���P���O�A�b��ӶʤƤ����L�{���A���̤@�����X�b�����ʦ��I�W�Ӥ��渨�C
�H�t�����]�l���һ��������ҡG�仲�]�l�N�c�T�a���X�b���ʤ��ߡA�ѻP�ʤƤ����C[46]�����Φ���������]�l�i�H�ѻP�ʤ�����٭������A�������X��ʤƦ��������������C
�ݭn���]�l���X�H�i��ʤƪ����A�b�����X���]�l�����p�U�A�Q�٬��滲�J���]apoenzyme�^�F�Ӧb���X�F���]�l��A�Q�٬������]holoenzyme�^�C�j�h�ƥ������A���]�l���O�H�D�@���s���覡�P�����X�F�]���@�Ǧ������]�l�i�H�P���@�����X�]�p��IJ�B�������J�C�IJ��i���^�C
���� NADH���Ŷ���R�����c�ҫ�
�����O�@���i�H�N�ƾǰ�αq�@�Ӗ��ಾ��t�@�Ӗ��W�������p���l�A�P�������P���a���X�A���S�w�������ʵo���O���n���C[45]���\�h���L�R�Ψ�l�ͪ��A�p�ֶ����B���i���M�����A���ݩ��C[47]�o�ǤƦX���L�k�ѤH��X���A�����q�L�����ɥR�C���P�����������a���ƾǰ�Τ]���P�GNAD��NADP+��a�B���l�A����A��a�A�Q��A������a���Q��AS-������J�i���]�i��a���Q��C[48]
�ѩ��b���ʤƤ�������ƾDzդ��o�ͤF�ܤơA�]���i�H�{�������O�@�دS��������Ϊ̺٬��u�ĤG����v�C�o�ةҿת��ĤG����i�H�Q�\�h���ҧQ�ΡC�Ҧp�A�ثe�w�������C�ʺؖ��i�H�Q�λ���NADH�i��ʤơC[49]
�b�ӭM���A�����᪺�����i�H�Q�A�͡A�H������M���@�צb�@��í�w�������W�C�Ҧp�ANADPH�i�H�q�L�C�ĥ��}�~�|�M�Ҳ��i�ĸ������ಾ���@�ΤU��S-������J�i�ĨӦA�͡C�ѩ����A��������������t��í�w�O���n���A�]���A�����A�ͨt����o�F�j�q������ǥH�Τu�~���ΡC[50]
[�s��] ���O��

�����εL���ʤƤ�����t�������i�{�P��q���Y�ϥܡC�i�H�ݥX�A�������S�������ʤƮɡA����q�`�ݭn��o���������Ư�~���F�L��A�A�M��~��ͦ������F�ӷ�������t�������ʤƮɡA�q�L����L��A��í�w�@�ΡA���C�F�F��L��A�һݯ�q�A�q�ӭ��C�F��Ӥ����һݪ���q�C
�P��L�ʤƾ��@�ˡA���ä����ܤ��������ű`���A�ӬO�q�L���C���������Ư��ӥ[�֤����t�v�]���k�ϡ^�C�q�`���p�U�A�����b���s�b�Τ��s�b����ر���U�A�������V�O�ۦP���A�u�O�e�̪������t�ק�֤@�ǡC���������X���O�A�b�����s�b�����p�U�A����i�H�q�L��L�����ʤƪ��u�ۥѡv�����ͦ����P�������A��]�O�o�Ǥ��P�������Φ��t�ק�֡C
���i�H�s����өΦh�Ӥ����A�]���i�H�Τ@�����O���W��e���o�ͪ������h�u�X�ʡv�t�@�Ӽ��O�ǤW���e���o�ͪ������C�Ҧp�A�ӭM�`�`�q�LATP�Q�����ѩҲ��ͪ���q���X�ʨ�L�ƾǤ����C
���i�H�P���a�ʤƥ��V�����M�f�V�����A�Өä����ܤ����ۨ����ƾǥ��šC�Ҧp�A�һ������i�H�ʤƦp�U��Ӥ��f�����A�ʤƭ��@�ؤ����h�O�̿��������@�סC[51]
 �]��´���F��CO2�@�פU�^ �]��´���F��CO2�@�פU�^ �]�����F�CCO2�@�פU�^ �]�����F�CCO2�@�פU�^
�@�@���������u*�v���ܡu�һ������v
���M�A�p�G�������ŷ��j�a�ͦV�_�Y�@��V�A��p����q�������A�Ӱf�������i��Ī��o�͡A�h���ɖ���ڤW�u�ʤƼ��O�ǤW���\����V�A�Ӥ��ʤƨ�f�����C
-

��@���誺���ʤƤ������z�G���]���ܬ��uE�v�^���X����]���ܬ��uS�v�^�A�óq�L�ʤƤ����ͦ������]���ܬ��uP�v�^�C
���ʤO�ǬO��s�����X�����O�M�ʤƤ����t�v����ǡC��s�̳q�L���������R�k�]enzyme assay�^����o�ΩʤO�Ǥ��R�������t�v�ƾڡC
1902�~�A���J�h�P��Q���X�F���ʤO�Ǫ��w�q�z�סF[52]�H��Ӳz�ױo��L�H�ҹ���X�i���̤��{�C[53]��Q�̤j�^�m�b��䭺�����X���ʤƤ����Ѩ�B�զ��G�����A����i�f�a���X�얡�W�A�Φ���-����ƦX���F�M��A������������ƾǤ������ʤơA������ͦ��������]�����ϡ^�C
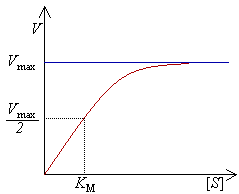
����l�����t�v�]���ܬ��u V�v�^�P����@�ס]���ܬ��u[S]�v�^�����Y���u�C�H�ۨ���@�פ��_�����A���������t�v�]�ͦV�_�̤j�����t�v�]���ܬ��u Vmax�v�^�C
���i�H�b�@�������ʤƼƦʸU�Ӥ����C�Ҧp�A�ŲM�Į֖�5'-�C�IJ��n���Ҷʤƪ������b�L�����p�U�A�ݭn�C�d�K�ʸU�~�~��N�@�b��������Ƭ������F�ӦP�˪������L�{�A�p�G�[�J�o�ز��n���A�h�ݭn���ɶ��u��25�@���C[54]�� �ʤƳt�v�̿���������M����@�סC�p�G�������s�b����N�J�ո��쪺�]���A�p���šB���ݪ�pH�M�����Q�@�סA���|�}�a�������ʡF�Ӵ���������t������ ��@�h�|�W�[�������ʡC�b���@�שT�w�����p�U�A�H�ۨ���@�ת����_�ɰ��A���ʤƪ������t�v�]���_�[�֨��ͦV�_�̤j�����t�v�]Vmax�^�]���k�Ϫ����M���u�^�C�X�{�o�ز{�H����]�O�A��������t�����誺�@�פɰ��A�V�ӶV�h�ۥѪ��A�U�������l���X����Φ���-����ƦX���F���Ҧ������l�����ʦ��I���Q���蹡�M���X�A�Y�Ҧ������l�Φ���-����ƦX���ɡA�ʤƪ������t�v�F��̤j�C���M�AVmax�ä��O���ߤ@���ʤO�DZ`�ơA�n�F��@�w�����t�v�һݪ�����@�פ]�O�@�ӭ��n���ʤO�ǫ��СC�o�@�ʤO�ǫ��ЧY�̤�`���]Km�^�A�����O�F��Vmax�Ȥ@�b�������t�v�һݪ�����@�ס]���k�ϡ^�C���S�w������A�C�@�ؖ�������S�xKm�ȡA���ܨ���P�����������X�j�ס]Km�ȶV�C�A���X�V�c�T�A�˩M�O�V���^�C�t�@�ӭ��n���ʤO�ǫ��ЬOkcat�A�w�q���@�Ӗ����ʦ��I�b�@�������ʤƨ��誺�ƶq�A�Ω���ܖ��ʤƯS�w���誺��O�C
�����ʤƮIJv�i�H��kcat/Km�ӿŶq�C�o�@���ܦ��S�Q�٬��S���ʱ`�ơA��]�t�F�ʤƤ������Ҧ��B�J�������`���C�ѩ�S���ʱ`�ƦP�ɤϬM�F������誺�˩M�O�M�ʤƯ�O�A�]���i�H�Ω������P�����S�w���誺 �ʤƮIJv�ΦP�@�ؖ���P���誺�ʤƮIJv�C�S���ʱ`�ƪ��z�׳̤j�ȡA�S�٬��X�������A����108��109 M-1s-1�F���ɡA���P���誺�C�@���I�����|�ɭP����Q�ʤơA�]���������ͦ��t�v���A�������t�v�ҥD�ɡA�Ӥ��l���X���t�v�_��F�M�w�ʧ@�ΡC�����o�دS�ʳQ�٬��u�ʤƧ������v�Ρu�ʤO�ǧ����ʡv�C�����������Ҥl���C�Ĥ��}���c���B�һ������B�A�Q�x�Pୖ��B�L��ƲB���B���J���Ė��B�]-���Q�i���M�W��ƪ��[�Ɩ��C
�̤��{�O�����q�@�Ωw���ӽT�ߪ��A�Ӹөw�߫h���ۥ��X���M���ʤO���X�ʪ��I���o�ǰ��w�C�M�ӡA�ѩ�/����/���������@�שM�ۤ����Ϊ̤@��/�G�����l�B�ʡA�\�h�ͤƩβӭM�i�{���㰾����q�@�Ωw�ߪ����w�C[55]�b�o�DZ��p�U�A�i�H���������̤��{�C[56][57][58][59]
?
�s�b�@�ǖ��A���̪��ʤƲ����ʤO�dzt�v�Ʀܰ�����l�X���t�v�A�o�ز{�H�L�k�Υ� �e���{���z�רӸ����C���h�زz�ҫ��Q���X�Ӹ����o���{�H�C�䤤�A�� �����p�i�H�Ζ�����誺���[�����Ӹ����A�Y�@�ǖ��Q�{���i�H�q�L�������q���Ӯ�������H�αN����H���T����\���ʤƬ��ʦ��I�C�t�@�زz�ҫ��ޤJ�F��� �q�l�z�ת�
���G�����A�Y��l�ιq�l�i�H��L�E�����S�]�N�p�P��L�G�D�@��^�A��������G�����٦����h��ij�C[60][61]�����ɵo�{���i����l�s�b�q�l���G�����C[62]�]���A����s�̬۫H�b���ʤƤ��]�s�b�۬��G�����A�i�H������L�������S�A�Ӥ��O���Dzβz�ҫ����覡�q�L���C���S�F��ʤƮĪG�C��������������ɴ��X�b�@���J��B�����ʤƤ������s�b���G�����A[63]�����G�����O�_�b���ʤƤ��������M�s�b�å����w�סC[64]
[�s��] ����@��
-
�����ʤƬ��ʥi�H�Q�h�ا���ҭ��C�C

���P�����������C�����ѦҦ� [65]�C�Ϥ��A�uE�v���ܖ��F�uI�v���ܧ���F�uS�v���ܨ���F�uP�v���ܲ����C
[�s��] �i�f����@��
�i�f����@�Ϊ��������h�ءA���̪��@�P�S�I�b��������ʪ���������㦳�i�f�ʡC
[�s��] �v���ʧ���@��
����P�����v�����X�������ʦ��I�]����M���褣��P�ɵ��X�쬡�ʦ��I�^�C����v���ʧ���@�ΡA�ʤƤ������̤j�����t�v�ȨS���ܡA���O�ݭn������@�סA�ϬM�b���[Km�Ȫ��W�[�C
[�s��] �D�v���ʧ���@��
�D�v���ʧ������i�H�P����P�ɵ��X�얡�W�A�Y��������X�쬡�ʦ��I�C��-����ƦX���]EI�^�Ζ�-���-����ƦX���]EIS�^���S���ʤƬ��ʡC�P�v���ʧ���@�άۤ�A�D�v���ʧ���@�Τ���q�L��������@�רӹF��һݤ����t�סA�Y���[�̤j�����t�vVmax�����ܤp�F�ӦP�ɡA�ѩ������v�T����P�������X�A�]��Km�ȫO�����ܡC
[�s��] ���v���ʧ���@��
���v���ʧ���@�Τ���֨��G�������P�B��ۥѪ��A�U�������X�A�ӥu��M��-����ƦX���]ES�^���X�A�b�������ʤO�ǤW���{��Vmax�MKm�ȳ��ܤp�C�o�ا���@�Υi��o�ͦb�h�Ȱ��C
[�s��] �ƦX����@��
�o�ا���@�λP�D�v���ʧ���@�Τ���ۦ��A�ϧO�b��EIS�ƦX���ݯd�������������ʡC�b�\�h�ͪ��餤�A�o������i�H�@���t���X������ �զ������C�Y�@�Ӗ���t�Ͳ��F�L�h�������A�����N�|����X���Ӳ���������t���Ĥ@�Ӗ������ʡA�o�N�i�H�O�Ҥ@���X�������h��������A�Ӳ������X���t�v �|�U���ΰ���C���o�ا���@�νձ������q�`���h�ȰA�è㦳�P�ձ��������X�����쵲�X���I�C�o�ا���@�Ϊ������t�v�P����@�ת����Y�Ϥ��A�O�����u�Φ� �OS�ΡC
[�s��] ���i�f����@��
���i�f����i�H�P�����X�Φ��@���s���A�Ө�L����@�Τ����P����������O�D�@�����X�C�o�ا���@�άO���i�f���A���@���Q�����N�L�k�A��_���ʪ��A�C�o������]�A�G�t�Ұ��i���]�@�إi�Ω�v���H���ξɭP�����ίg���Ī�[66]�^�B�f�Ұ��D�Q�t�]PMSF�^�B�C�`���M���q�ǪL�C�o���Ī����O�P�����ʦ��I���X�ֳQ�E���A�M��P���ʦ��I�B���@�өΦh���i��Ĵݰ�o�ͤ��i�f�������Φ��@���s���C
[�s��] ������γ~
������`�Q�Χ@�Ī��A�P�ˤ]�i�H�Q�@���r�ĨϥΡC���Ī��M�r�Ĥ������t�O�q�`�D�`�p�A�j�h�ƪ��Ī������@�w�{�ת��r�ʡA���p���Զ뺸Ĭ���Ҩ��G�u�Ҧ��F�賣���r�A�S������O�L�r���v�]�uIn all things there is a poison, and there is nothing without a poison�v�^�C[67]�ۦP���A�ܥͯ��M��L�ܷP�V�Ī��u�O�S���ʦa���f�����Ӥ��O���J�D���r�ʡC
�@����o�s�x���Ϊ�����Ī��O���q�ǪL�A���i�H�������[�������ʡA�����[�i�H�Ͳ����g�����H���e�C�����A�]���A���q�ǪL�i�H�_�����k�h�P���g���@�ΡC�Ӽ@�r�r����ƪ��i�H�q�L���X�ӭM�����Ɩ����I�B���ɩM�K��l���i�f�a������ʡA�q�ӧ���ӭM���I�l�@���C[68]
[�s��] ���ʱ���

�Ѩ�ӨȰ�զ��� �y�P�f�r�����J�� ����y���������l���T�����c�ϤΨ�P�ӭM�����X�ܷN�ϡC��ܬ��Ŧ⪺�Ȱ�i�H�S���ʦa���X�H��ӭM�������}��F�b�f�r���X��ӭM��A��ܬ����⪺�Ȱ�i�H�ҰʫI�V�{�ǡC
�ӭM�������ر���ʤƬ��ʪ�����G
- �ھڥ~�����Ҫ��ܤơA�ӭM�i�H�W�j�δ�z�����Ͳ��]�Y��������]������M��Ķ�^�C�o�ݩ�@����]�ձ��A�Q�٬��������ɩM�����C�Ҧp�A�����Ҥ��X�{�p�C�`���o�˪��ܥͯ��ɡA�����ӵߥi�H���ܥͯ����ͧܩʡA���]�N�b��ӵ��餺���]-�b�ſ}�����Q���ɦӤj�q�Ͳ��A�o�ؖ��i�H���ѫC�`�����l�W���䪺�]-���i���C�t�@�ӨҤl�O�b�H���xŦ���s�b�@��������Ī��N���D�`���n�����A�ӭM���P450��Ɩ��F��o�@���������ɩΧ���A�|�ɭP�Ī��ۤ��@���C
- �q�L�b���P���ӭM�դ����i�椣�P���N�³~�|�A���i�H�Q���j�C�Ҧp�A�תջ����X���O���ӭM�����B������M�������������@�t�C���ҧ����A�ӯתջĪ����ѡ]�H���ѯ�q�^�O�b�ɽu�餤�ѥt�@�t�C���q�L�]-����ӧ����C[69]
- ���i�H�Q����P���ƾ��ҽձ��C�Ҧp�A�@�ӥN�³~�|�����ײ����`�`�O�o�@�~�|���Ĥ@�Ӗ�������A�q�ӽձ��o�@�N�³~�|�������q�C�o�ؽձ�����Q�٬��t���X�����A�]���ײ������X���q�O����ۨ��@�ձ��C�t���X����i�H�ھڲӭM���ݭn�A���Ħa�ո`�����N�ª����X���t�v�A�q�ӨϲӭM����q�M���誺���t���ġA�è���h�l�������X���C������@�ΡA�i�H�b�ͪ��餺�����@��í�w���������ҡ]�Y�餺�����^�C
- ��Ķ����]�i�H�ձ��������ʡC�o�ǭ��]�A�C�Ĥ��B�ר��M���ƩM�}����C�Ҧp�A�ӭM�����خq���H����A��]�A�}��X�����b�����h�Ӗ��i���C�ĤơA���U�����}�����X���έ��ѡA�ϱo�ӭM�i�H����}���ܤƲ��ͤ����C[70]�t�@����Ķ������Ҥl�O�h���쪺�Ť��C�ؾ��ųJ�Ֆ��A�@�خ��Ʃ��J�Ֆ��A�O���ͩ���Ŧ�����L���ʪ��ؾ��ųJ�Ֆ����A�o�@�J�ճq�L�B���F�G��~�Q�E���C�o�ؤ覡���Ħa����F�ؾ��ųJ�Ֆ��b�i�J�z���e���Ư�Ŧ�Ψ�L��´�C�o�صL���ʪ������e��Q�R�W�������C
- �٦��@�ǖ��i�H�q�L�w��줣�P���ҫ�ӳQ�E���A��p�q�٭�A�����ҡ]�ӭM���^���ƺA���ҡ]�ӭM�P��Ŷ��^�A�q��pH���Ҩ�CpH���ҵ��C�y�P�f�r������y�������J���N�O�@�ӨҤl�G������IJ��J�D�ӭM�n�w���ĩ����ҮɡA�����c�H�ߨ�o���ܤơA�ɭP����o�E���C[71]
[�s��] �����e�f
�������ʥ����Y�汱��H�����餺�����A������v�T�@�����䖡���\������]�ʳ��]�p�����ɭP�����ܤơA�L�q���F�B�L�C���F�ΧR�����ܡ^���i��ɭP��ǩʯe�f�o�͡C�\�h�ƹ���ܡA�@�حP�R�e�f���f�]�i�H�u�O�ѩ�H�餤���Ƥd�ؖ������@�صo�ͥ\��G�١C
- �f��ଧ��g�G���دf�g�O�嫬���������f�Ҥ��@�C�f�]�O�f���i���m�Ɩ��]��\��O�ʤ��f���i�����ѹL�{�����Ĥ@�B�^�W�@���i��Ħ��I�o�ͤF���ܡA�ɭP�餺�f���i���M���������������L���A�p�G�S���o��X�A���v���A�|�i�@�B�ɭP�����ê�C
- �v���f�G�ӯf�O�ѩ�������ͪ��X���~�|���S�w���������ʹL�C�]��]���ܩΨ�L��]�ɭP�^�A�ϱo���������v�������ͩM�ƪn���`�A�b�@�w���]�]�p�����Ӯg�^�U�A�i�ɭP�ֽ��Ψ�L��´���x�o�ͯf�ܡC
- ���ʹӭM���s�XDNA�״_����������]�o�ͬ��ܡA�䵲�G�|�ɭP��ǩ����g��X�f�x�A�p�ۦ�ʤz�֯g�CDNA�״_�����ʳ��ɭP�H��ॢ�״_���ܰ�]����O�C�o�ͪ����ܤ��_�n�֡A�̲רϱo�w�̦��h�����g�o�͡C
[�s��] �R�W�W�h
���q�`�O�ھڨ����ʽ�Ψ�Ҷʤƪ��ƾǤ��������өR�W�]�^�夤�A�b������̫�n�[�W-ase�����^�A�p�ſ}���]lactase�^�B�J��B���]alcohol dehydrogenase�^�MDNA�E�X���]DNA polymerase�^�C���o�˩����|�ɭP�㦳�ۦP�\����P�����]�P�u���^���ۦP���W�r�A�ӦP�u�����ۤ��P���i��ħǦC�åi�H�q�L���̤��P���̾ApH�M���P�������ʤO�ǰѼƨӰϤ��C�ӥB�A�@�ǖ��㦳��өΦh�������ʤƬ��ʡA�o�ةR�W�覡�N�|�ϱo�P�@�ؖ��㦳���P���W�r�C���F�ѨM�o�ǰ��D�A��ڥͪ��ƾǻP���l�ͪ����p���o�i�X�F�����t�ΩR�W�k�G
EC�s���W�w�C�ؖ����ѥ|�ӼƦr�Ӫ��ܡA�æb�Ʀr�e�a�H�uEC�v�C�䤤�A�Ĥ@�ӼƦr�O�ھ����P�������ʽ�ӱN���j�P�������j���G
��ڨt�Τ����k�����W�z�����N���̦��s���~�A�ٮھږ��Ҷʤƪ��ƾ������S�I�M�ѥ[��������Ϊ����P�A�N�C�@�j���S�i�@�B�����C�s�������Ĥ@�ӼƦr���ܸӖ��ݩj���������@���F�ĤG�ӼƦr���ܸӖ��ݩ���@�����F�ĤT�ӼƦr���ܨ�-�����F�ĥ|�ӼƦr�O�Ӗ��b��-���������ƧǡC
���Q�Ω�Ƥu���U���ݭn���ׯS���ʶʤƱ��p���γ~�C���O�A���q�`����ʤƪ������ƶq�����A�ӥB���̦b�L�����G���M���ű��p�U�ʥFí�w�ʡC���F�����������ΩʡA�Q���J�ս�u�{�q�L�X�z�]�p����~�i�ƨӳy�X�㦳�s�S�I�]�Ҧp�@���š^�����w�g�����@�Ӭ��D����s���C[72][73]�o����s�u�@�]���F���\���Ҥl�A�@�ǯ���ʤƦ۵M�ɤ������ҵL�k�ʤƪ����������w�g�}�l�Q�]�p�X�ӡC[74]
�H�W�峹�Ӧ۩����ʬ�A���\Ū�����I�H�U�s��
http://zh.wikipedia.org/zh-tw/%E9%85%B5%E7%B4%A0
�^�W�@��
���� Twitter!
���� Plurk!
���� Facebook!
|
|
|